By Andrew L. Rypel
“The Delta is full of species that thrive in the lakes in southern Arkansas” ~Bill Bennett

by Andrew Rypel
As we rapidly enter another drought, long-standing questions on ecological impacts of increased temperatures, reduced water levels and flows re-emerge. This reality recently reminded me of some of my own previous work looking at growth rate variations of largemouth bass in response to droughts in the southeastern USA (Rypel et al. 2009). Results from this work may be useful/interesting for biologists and managers in California considering similar questions.
While droughts elsewhere occur, they do so on different time tables and climatic cycles than in CA. Yet many of the species and underlying ecological dynamics remain similar (Marchetti et al. 2001, Scott and Helfman 2001, Rahel 2002, Moyle & Mount 2007). The homogenization of ecological communities on Earth is one of our greatest challenges, and parallels human modifications to landscapes, cities and food systems. For example, when traveling to far flung destinations, one could with little effort, eat at all the same restaurants, shop at the same stores, and access almost the exact same products. Similarly, most aquatic systems with suitable temperatures and fish habitat across North America (and other continents!) contain species like bluegill, largemouth bass, common carp and so on. Today, we’ll focus on largemouth bass, a species common throughout California. Indeed it is important to remind ourselves that California shares a non-native fish fauna with other states, that we can learn from, and that warm-water fishes are expanding in CA. California is home to some of the largest black bass captured on Earth, and there is some evidence that they compete with and prey on our rapidly declining native fishes. So, they’re definitely worth thinking about.

During the mid-2000s, I was assisting the Georgia Department of Natural Resources by researching largemouth bass populations from some of their reservoirs. Biologists had been extracting otoliths from bass populations to calculate and study the usual metrics: growth mortality, age class diversity and reproductive success. Otoliths are calcified bony structures in the inner ear of fishes. They provide fishes a sense of equilibrium in the water, and lay down concentric rings, much like tree-rings. I had similar samples from ecosystems in Alabama and Mississippi.
Bass fisheries are the primary focus for many fisheries managers in the southeast, and generate extraordinary economic impact locally (Chen et al. 2003). Bass fisheries are also important in California, but receive somewhat less adulation than in southeast.
During this work (circa 2006-2008), a historic drought struck the region, and reservoir levels plunged. This quickly led to concern among anglers and the public over what might be happening with fish populations. Coincidentally, I was also exploring dendrochronology (tree-ring) techniques applied to fish otoliths, and the idea was hatched to measure incremental growth of each fish in all of its years of life. Using this approach, we might better understand conditions when bass growth well or poorly, including how bass responded to drought, specifically.
I examined 397 bass otoliths from 13 waterbodies across Georgia, Alabama and Mississippi. Using the annual increments led to a much larger sample of 1450 annular growth increments. Each growth history for each fish was detrended to remove growth trends related to fish growing more when they are younger and less when older. Interseries correlations were high, ranging 0.27-0.88, with a mean of 0.62. This result indicated that, within populations, individual bass growth was highly synchronous over time among individuals. It was also immediately clear that the years 2000 and 2007 (two major drought years) were “pointer years” – with exceptionally high growth across populations. In contrast, 1994, 1997, and 2003 were usually poor years of growth.
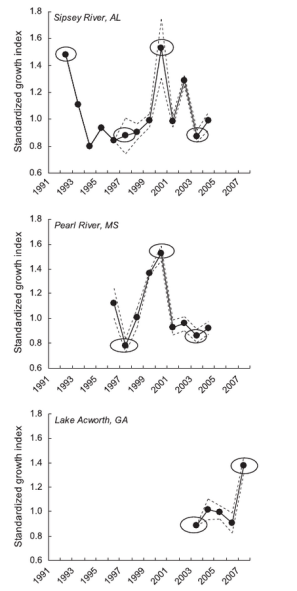
When growth rates were correlated with various climate indices, the pattern became clear: Bass growth tended to increase with temperature and decrease with precipitation. Droughts appeared to be good for bass. In fact, the pattern was so strong that ~50% of bass growth in any ecosystem could be predicted based solely on climate variables. I found this statistic to be astounding, especially given that density-dependent effects (e.g., growing less when population sizes are large) in bass populations can be quite strong. However, a related study clearly indicated that lipid (fat) deposits in seven warmwater species and thousands of fish in an Alabama Reservoir spiked up to 5x during the 2000 drought (Rypel and Bayne 2009).
There are a few potential explanations for the pattern. Most importantly, largemouth bass are a warmwater fish species. As such, their metabolism and vital rates increase directly as a function of temperature, up to some very high thresholds (Glover et al. 2012), especially compared to California’s cold-adapted native fish fauna (Jobling 1981, Zillig et al. 2021). Second, reduced water levels may concentrate prey like sunfishes and shad (Schlosser 1985, Craven et al. 2010). Finally, reduced water levels may also stimulate nutrient cycling and primary production. For example, large algal blooms are often observed in reservoirs during unusually hot and dry summers (Braga et al. 2015), which can temporarily boost food availability for small fish.
So unfortunately, increased temperatures and droughts are likely boons for non-native warmwater non-native species like largemouth bass. It is worth noting that this study was done on the other side of the country; thus we don’t know if these results translate to the arid West and California. However, black basses have been on the march for some time (Rypel et al 2017). In the upper midwest, largemouth bass populations have increased dramatically, while populations of native cool- and cold-water fishes like Walleye and Cisco have declined (Sharma et al. 2011, Rypel et al. 2018). There is no reason these patterns should be different in California ecosystems. Indeed they may even be more pronounced because the benefits to basses during droughts will likely assist non-native populations further expand their footprint. A statewide assessment of climate change effects on native and non-native fishes indicates that warmwater fishes such as largemouth bass will benefit from increased temperatures and lower reservoirs, while most native fishes will have their declines hastened (Moyle et al. 2013).
The future is now, and it looks like it is going to involve large and fast-growing black bass populations.

Further Reading
Braga, G. G., V. Becker, J. N. P. d. Oliveira, J. R. d. Mendonça Junior, A. F. d. M. Bezerra, L. M. Torres, Â. M. F. Galvão, and A. Mattos. 2015. Influence of extended drought on water quality in tropical reservoirs in a semiarid region. Acta Limnologica Brasiliensia 27(1):15-23.
Chen, R., K. Hunt, and R. Ditton. 2003. Estimating the economic impacts of a trophy largemouth bass fishery: issues and applications. North American Journal of Fisheries Management 23(3):835-844.
Craven, S. W., J. T. Peterson, M. C. Freeman, T. J. Kwak, and E. Irwin. 2010. Modeling the relations between flow regime components, species traits, and spawning success of fishes in warmwater streams. Environmental Management 46(2):181-194.
Gaeta, J. W., G. G. Sass, and S. R. Carpenter. 2014. Drought-driven lake level decline: effects on coarse woody habitat and fishes. Canadian Journal of Fisheries and Aquatic Sciences 71(2):315-325.
Glover, D. C., D. R. DeVries, and R. A. Wright. 2012. Effects of temperature, salinity and body size on routine metabolism of coastal largemouth bass Micropterus salmoides. Journal of Fish Biology 81(5):1463-1478.
Jobling, M. 1981. Temperature tolerance and the final preferendum—rapid methods for the assessment of optimum growth temperatures. Journal of Fish Biology 19(4):439-455.
Marchetti, M. P., T. Light, J. Feliciano, T. Armstrong, Z. Hogan, and P. B. Moyle. 2001. Homogenization of California’s fish fauna through abiotic change. Pages 269-288 in J.L. Lockwood and M.L. McKinney, editors. Biotic Homogenization. Kluwer/Academic Press, New York.
Moyle, P.B. and J. Mount. 2007. Homogenous rivers, homogenous faunas. Proceedings of the National Academy of Sciences 104: 5711-5712.
Moyle, P.B., J. D. Kiernan, P. K. Crain, and R. M. Quiñones. 2013. Climate change vulnerability of native and alien freshwater fishes of California: a systematic assessment approach. PLoS ONE 8(5): e63883.
Rahel, F. J. 2002. Homogenization of freshwater faunas. Annual Review of Ecology and Systematics 33(1):291-315.
Rypel, A. L. 2009. Climate–growth relationships for largemouth bass (Micropterus salmoides) across three southeastern USA states. Ecology of Freshwater Fish 18(4):620-628.
Rypel, A. L., and D. R. Bayne. 2009. Hydrologic habitat preferences of select southeastern USA fishes resilient to river ecosystem fragmentation. Ecohydrology 2(4):419-427.
Rypel, A. L., D. Goto, G. G. Sass, and M. J. Vander Zanden. 2018. Eroding productivity of walleye populations in northern Wisconsin lakes. Canadian Journal of Fisheries and Aquatic Sciences 75(12):2291-2301.
Rypel, A. L., J. Lyons, J. D. T. Griffin, and T. D. Simonson. 2016. Seventy-year retrospective on size-structure changes in the recreational fisheries of Wisconsin. Fisheries 41(5):230-243.
Schlosser, I. J. 1985. Flow regime, juvenile abundance, and the assemblage structure of stream fishes. Ecology 66(5):1484-1490.
Scott, M. C., and G. S. Helfman. 2001. Native invasions, homogenization, and the mismeasure of integrity of fish assemblages. Fisheries 26(11):6-15.
Sharma, S., M. J. Vander Zanden, J. J. Magnuson, and J. Lyons. 2011. Comparing climate change and species invasions as drivers of coldwater fish population extirpations. Plos One 6(8):e22906.
Zillig, K. W., R. A. Lusardi, P. B. Moyle, and N. A. Fangue. 2021. One size does not fit all: variation in thermal eco-physiology among Pacific salmonids. Reviews in Fish Biology and Fisheries 31:95-114.
Andrew Rypel is a professor of Wildlife, Fish, and Conservation Biology and Co-Director of the Center for Watershed Sciences at the University of California, Davis.
Leave a Reply