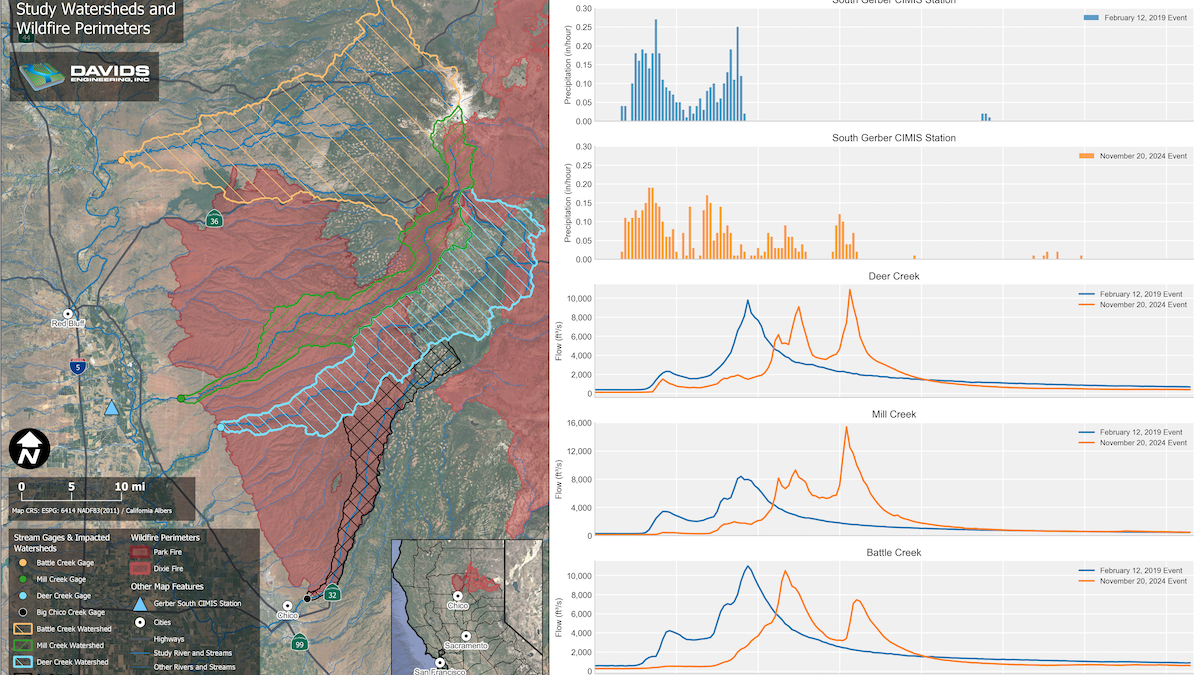
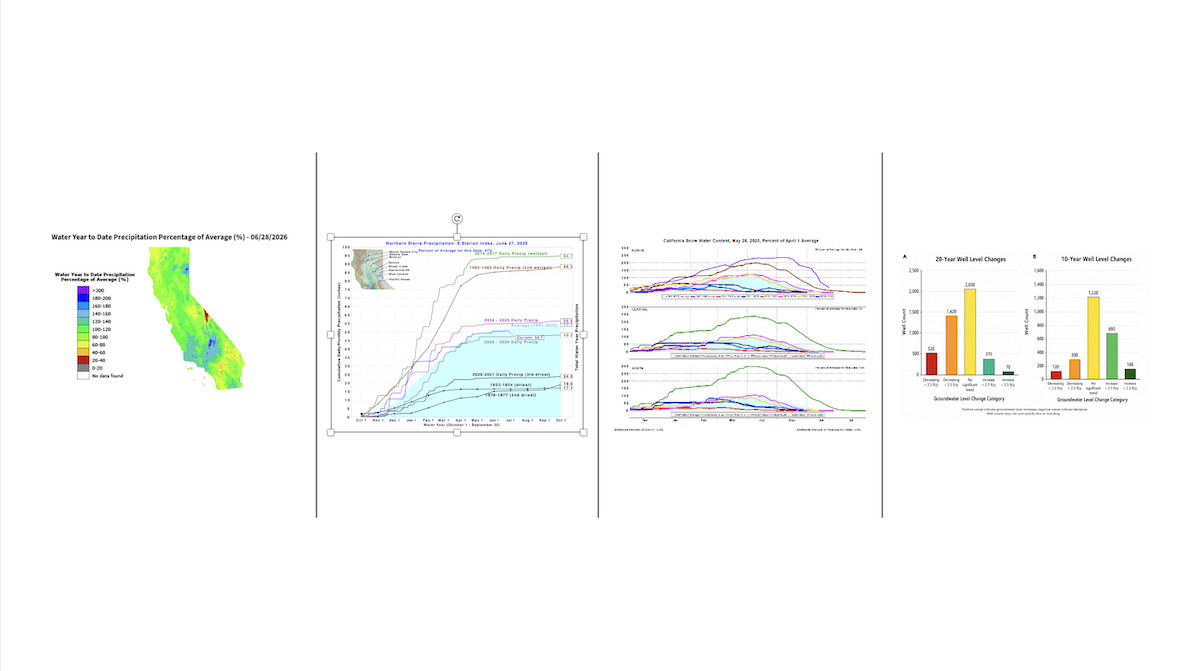
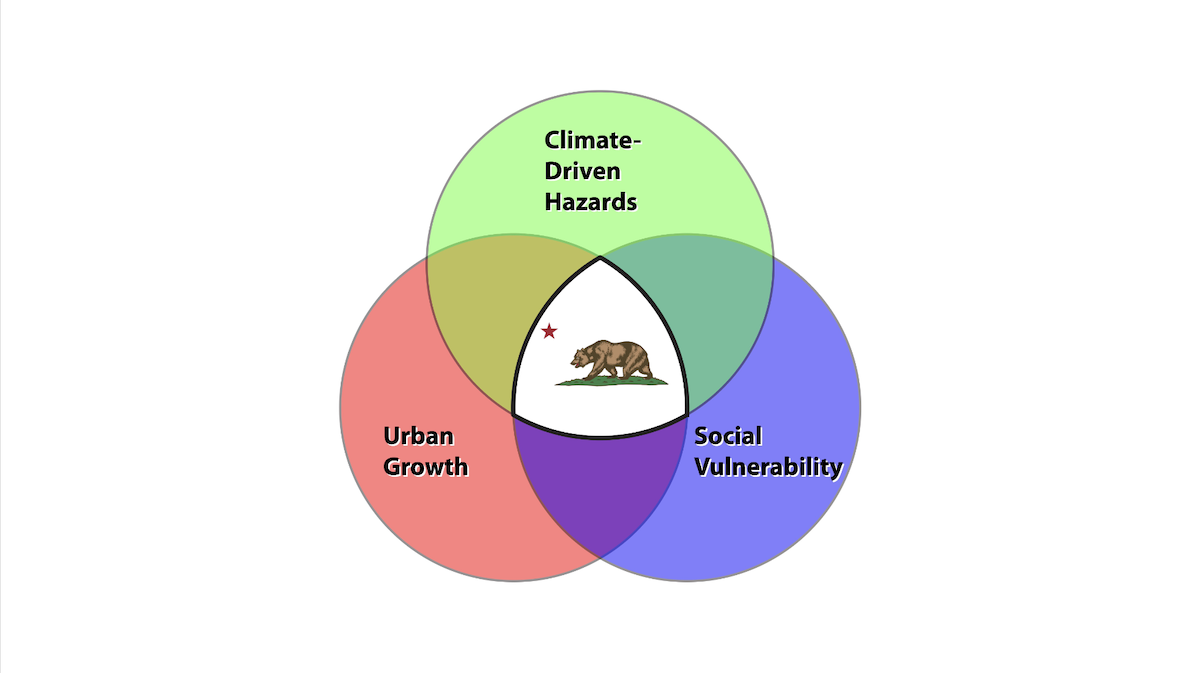
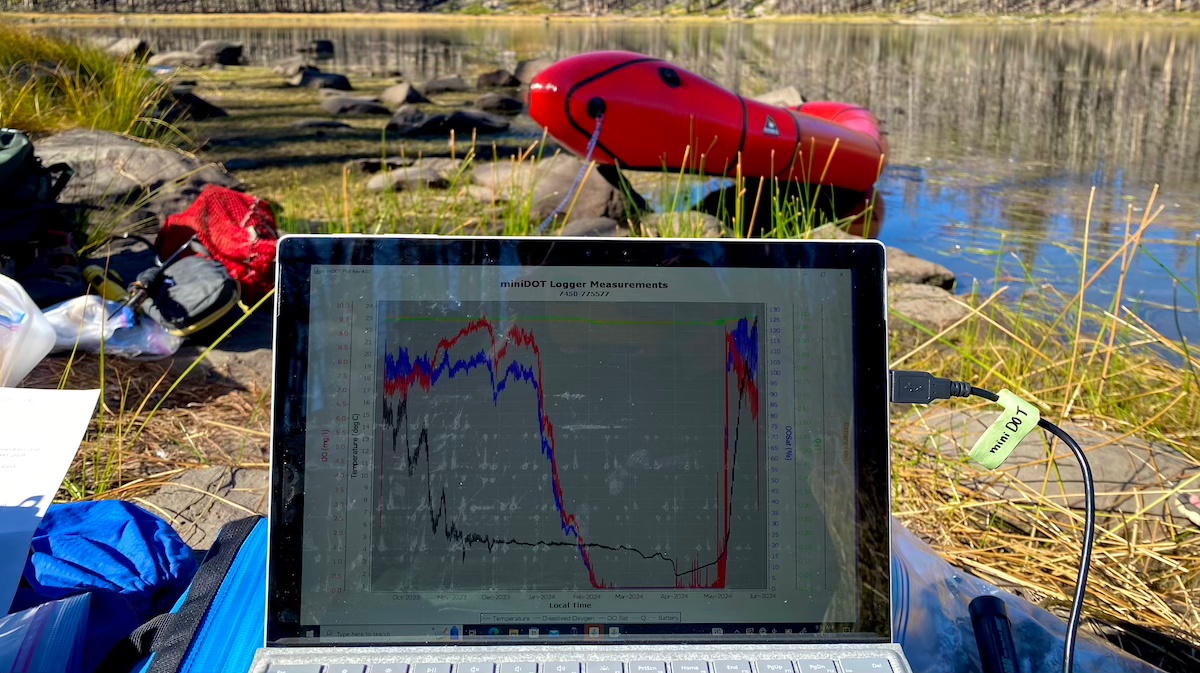
Latest
-

Where are they now: Francine De Castro
By Francine De Castro . . . I worked in the Aquatic Research Collective (ARC) lab with Dr. John Durand from 2019–2021 as an undergrad student, and my duties mostly included zooplankton ID with the occasional foray into the field to assist with the long-term Suisun and Arc sampling projects. My favorite memory as a…